
MAFLD under the lens: the role of gut microbiota


Abstract
Obesity, the metabolic syndrome, and metabolic dysfunction-associated fatty liver disease (MAFLD) can be portrayed as transmissible diseases. Indeed, they can be induced, in animal models, by cohabitation or by transplantation of fecal microbiota from other animals or humans with those diseases. As such, to get a 10,000-foot view, we need to see under the lens the microbes that populate our gut. Gut microbiota participates in the harvesting of energy from nutrients, it allows the digestion of otherwise indigestible nutrients such as fibers, and it also produces short chain fatty acids and some vitamins while emitting different compounds that can regulate whole-body metabolism and elicit proinflammatory responses. The metabolic syndrome and MAFLD share physiopathology and also patterns of gut dysbiota. Moreover, MAFLD also correlates with dysbiota patterns that are associated with direct steatogenic or fibrogenic effects. In the last decade, a tremendous effort has allowed a fair understanding of the dysbiota patterns associated with MAFLD. More recently, research is moving towards the delineation of microbiota-targeted therapies to manage metabolic dysfunction and MAFLD. This review provides in-depth insight into the state-of-the-art of gut dysbiosis in MAFLD, targeting clinical hepatologists.
Keywords
INTRODUCTION
The steady increase in the prevalence of metabolic-associated fatty liver disease (MAFLD) since its description in the 1980s has turned it into a major health threat[1]. Indeed, MAFLD was the fastest growing indication for liver transplantation over the last 20 years and is already the leading cause for liver transplantation among women and patients over 54 years in the US[2].
MAFLD is the hepatic expression of metabolic dysfunction and goes hand in hand with obesity and insulin resistance (IR)/type 2 diabetes mellitus (T2DM)[3]. Its pathogenesis is complex and dependent on additional individual susceptibility, whether genetic, behavioral, or acquired[4]. The gut microbiota is one such factor that helps explain individual susceptibility to MAFLD and liver disease progression[5]. In the last decade, we witnessed a research boom on the role of gut dysbiota in MAFLD, regarding its pathogenesis, diagnostic ability, and role as a therapeutic target.
The way the gut microbiota is assessed has been changing since the first studies before the 1990s, when only culture-based methods were available[6]. Even though culture-based methods are cheap, less than 30% of the gut microbiota has been cultured. With the development of culture-independent techniques, there has been an exponential growth in the knowledge of gut microbiota. These techniques have shown the diversity of the gut microbiota, allowing quantitative and/or qualitative information about bacterial species and in-depth comparison of the microbiota from healthy and MAFLD patients[7].
Most studies used culture-independent, biomarker-based profiling techniques that involve sequencing a ubiquitous bacterial gene - small subunit ribosomal RNA (16S rRNA) - which is highly conserved but divergent enough to allow resolution at a genus level and, in some cases, at the level of species or even strains[8]. This provides a relatively accurate fingerprint of microbial community composition but cannot accurately identify bacteria on a strain level, nor does it reflect the microbiota functional properties, even though it is possible, through algorithms, to infer in silico metagenomic analyses from 16S RNA data[9].
With recent advances in computational biology and high-throughput sequencing technology (shotgun sequencing or pyrosequencing), research is moving to untargeted whole genome sequencing and “omics” approaches[6]. They include metagenomics (determination of the functional genes encoded), metatranscriptomics (determination of the functional genes expressed), metaproteomics (identification of proteins), and metabolomics (identification of bioactive small molecules)[8,10,11]. Shotgun sequencing metagenomic data allow the characterization of the DNA library from a microbial community with an accurate portrayal of the potential microbial functional properties. However, it does not inform the activity or directionality of a certain present pathway. Those are better inferred by metatranscriptomics and metabolomics[12].
The interpretation and extrapolation of the studies of gut microbiota are challenging. First, the microbiota is as specific to an individual as a fingerprint[13]; that is, each person has a unique collection of bacterial strains and species, although there is the controversial concept of the existence of a core functional microbiome[14,15]. There is also appreciable intraindividual variability with age, medication, and changes of lifestyle[8]. At a population level, the gut microbiota is modulated by several factors, such as geographic localization, different dietary patterns, host genetics, and environmental factors. Most studies did not account for confounding factors that are known to influence the microbiota composition, which may influence the conclusions and the extrapolation of the results to all patients with the same condition. Besides bacteria, gut microbiota also has other communities, such as viral and fungal, but their role in the crosstalk between microbes and host is even less known[8,12]. Sampling may represent another weakness. To start, the choice of the sample will render different results, since the microbiota from stool (representing luminal content) will be different from those from mucosal biopsy samples. The way stool is collected, stored, and processed can have a major impact on the quality of the samples, since it may allow contamination and a variable degree of proliferation or death of different bacteria that will change the relative proportions of bacterial species[7]. Taking everything into consideration, it is understandable why there are striking differences between studies, which hamper the generalization of the results.
We present an in-depth review of the state-of-the-art of gut microbiota in MAFLD with basilar concepts directed to clinical hepatologists.
OVERVIEW OF THE GUT MICROBIOTA
The term gut microbiota refers to all microorganisms found in the digestive tract. It is considered an essential invisible organ[16], an example of a dynamic and complex symbiotic relationship between host and microbiota that mutually influence each other[17]. The gut microbiota is composed of bacteria, archaea, fungi, and viruses. The most represented bacteria phyla are Bacteroidetes, Firmicutes, Actinobacteria, and Proteobacteria. Three enterotypes are described according to the bacterial genus predominance: Bacteroides in type 1, Prevotella in type 2, and Ruminococcus in type 3[15].
The term microbiome refers to the combination of genomes and genes from the members of a microbiota[18]. The microbiome acts as a massive functional booster of the host genome, harboring more than 100-fold the number of human genes[19]. Indeed, these extra genes encode enzymes that are not encoded by the host, adding functions that would otherwise be absent in humans, such as the breakdown of complex polysaccharides and the synthesis of polyphenols, essential amino acids, and vitamins[19-21]. There is great functional redundancy across taxa, and changes in microbiota composition may not result in functional consequences[22].
Dysbiosis refers to the permanent disruption of the symbiotic relationship between host and microbiota, with changes in the composition and function of the microbiota. Dysbiosis has a profound impact on host physiology and may promote the development of diseases such as obesity, T2DM, and MAFLD[23-25]. A ubiquitous feature of dysbiosis is the loss of species diversity, which is counterbalanced by the overgrowth of proinflammatory species, promoting intestinal inflammation[23]. Alpha-diversity refers to the bacterial richness in one sample and beta-diversity to the variation in a group of samples[26].
The gut microbiota has several functions with potential benefit for the host homeostasis: maintenance of mucosal barrier integrity; defense against pathogenic microbes; and nutrients, bile acids, and drug metabolism[25].
The interaction between enterocytes and gut microbes regulates epithelial permeability[27]. For example, the commensal Akkermansia muciniphila enhances tight junctions’ function[28,29]. Consequently, dysbiosis is frequently associated with a leaky gut[23]. Gut microbiota further contributes to defense by constraining pathogens colonization through the competition for attachment sites and the production of bacteriocins that inhibit their competitor’s growth[30]. Consequently, in dysbiosis, there is a shift in this balance allowing pathogenic strains to overgrow. Importantly, the gut microbiota is also a key regulator of innate and adaptive immunity[31], acting on gut-associated lymphoid tissues, regulatory T cells, IgA producing plasma cells, innate lymphoid cells, resident macrophages, and dendritic cells in the lamina propria[32].
Regarding nutrient metabolism, the gut microbiota acts on the metabolism of carbohydrates, vitamins, and amino acids. Colonic microbes ferment complex carbohydrates producing volatile organic compounds and short-chain fatty acids (SCFA), mostly propionate, butyrate, and acetate[33]. The critical physiological functions of SCFA are as follows: maintenance of intestinal mucosa integrity, a source of energy for the host, modulation of glucose and lipid metabolism, control of energy expenditure, and immune regulation[34-36]. Butyrate is the most crucial SCFA for human health, being the major source of energy for colonocytes, presenting anti-colon cancer activity, and potentially decreasing gut permeability[37,38]. Propionate may abrogate hepatic gluconeogenesis and promote satiety, with the potential to reduce adiposity. Acetate is the most abundant SCFA and modulates gut microbiota, for example, being an essential metabolite for Faecalibacterium prausnitzii growth[39].
The gut microbiota is responsible for the synthesis of vitamins K and B (including biotin, cobalamin, folates, nicotinic acid, panthotenic acid, pyridoxine, riboflavin, and thiamine)[39,40]. It can also extensively degrade proteins from diet, host enzymes, mucin, and sloughed-off intestinal cells, producing amino acids and their derivatives, including IR-linked branched-chain amino acids (BCAA)[41,42].
The metabolism of bile acids results from the interactive crosstalk between the host and the gut microbiome. Primary bile acids (cholic acid (CA) and chenodeoxycholic acid (CDCA)) are cholesterol-derived metabolites, synthesized and conjugated with glycine or taurine in hepatocytes[43]. They are mostly absorbed by active transport in the terminal ileum, entering the enterohepatic circulation. The gut microbiota metabolizes (deconjugation, 7-dehydroxylation, and 7-dehydrogenation) them into secondary bile acids [deoxycholic acid (DCA) and lithocholic acid (LCA)], changing their structure and function[44]. There is a bidirectional relationship between gut microbiota and bile acids. Bile acids have selective antimicrobial characteristics that modulate the gut microbiota composition[43]. Besides the role of bile acids in fat absorption, they act through different receptors [e.g., farnesoid-X receptor (FXR) and Takeda-G-protein receptor (TGR5)] as well as receptor-independent mechanisms such as modulating membrane dynamics[45]. Bile acids modulate glucose metabolism (e.g., promote hepatic glycogen synthesis and insulin sensitivity and increase pancreatic insulin secretion) and energy homeostasis (facilitate energy expenditure, favor thermogenesis, and mediate satiety in the brain)[46], but they can also induce hepatic lipogenesis, cell injury, and proinflammatory and profibrogenic responses[22].
The microbiota composition varies along the digestive tract, with different strains in the mouth, small intestine, and different colorectal localizations, as well as transversal variations (gut lumen, mucous layer, and intestine villous/cripta)[47].
Several factors regulate the microbiota composition, such as environmental factors (delivery mode, diet, antibiotics, and prebiotics/probiotics), host factors (genetic), environmental factors, and inter-microbial interaction (competition between strains).
Acute perturbations of the gut microbiota, such as acute diarrhea, short-term diet manipulations, or short-term antibiotic use, induce only transient modifications that tend to return to baseline over time, a characteristic known as resilience. However, if the perturbation perpetuates in time (e.g., long-term dietary manipulations), it can overcome microbiota resilience and lead to sustained changes in the gut microbiota[10,48,49].
The newborn microbiota is transferred vertically and is influenced by the delivery route: either colonized by microbes from the maternal vagina or from maternal skin flora with cesarean section birth[50,51]. Microbiota changes with age, increasing its diversity from childhood to adulthood and then decreasing in the elderly[23].
Diet seems to shape gut microbiota in infants (breast milk or formula) and adulthood. For example, preclinical and epidemiological studies showed an association between high-fiber diets rich in fruits and vegetables and higher gut microbiota diversity and richness[32], as well as decreased Firmicutes/Bacteroidetes ratio, increased levels of Prevotella and Xylanibacter, and a decrease in proinflammatory pathogens Enterobacteriaceae, Shigella, and Escherichia. On the contrary, the high-fat, high-protein Western diet is associated with a decrease in diversity and an increase in Firmicutes[48,52-54].
Geographic localization is also a strong determiner of gut microbiota composition. Indeed, a study evaluating individuals from four districts within one province of China found that host location showed the strongest association with microbiota variation[55]. Furthermore, in developed countries, there is a trend of a loss of microbiota diversity. Geographic variations might be the result of different modulators of the gut microbiota, such as diet, lifestyle, sanitary conditions, proportion of infants delivered by cesarean sections, use of antibiotics, and genetics[56,57].
Studies with twins showed that host genetics is significantly associated with the abundance of specific gut microbial taxa, dubbed hereditable taxons. For example, there seems to be an association between Bifidobacteriaceae and the LCT locus (higher abundance of Bifidobacterium in lactose-intolerant subjects that maintain a regular intake of dairy products)[58-60].
Longitudinal studies showed that the microbial composition of a unique host may change in function of diet, drug intake, lifestyle (smoking, travelling, and physical activity), co-morbidities, and colonic transit time[10,49].
GUT MICROBIOTA AND OBESITY
The pathophysiology of obesity is complex and does not reduce to a simple arithmetic between the energy consumed and expended. Behavioral, genetic, and environmental factors concur.
The first evidence for the role of gut microbiota in obesity and the concept of obesity as a transmissible disease comes from seminal preclinical studies from the beginning of the century[61]. Those studies showed that rodents devoid of microbiota were resistant to diet-induced obesity and would adopt an obese phenotype when submitted to fecal microbiota transplantation (FMT) from genetically or diet-induced obese rodents but not from lean donors[62-64]. Furthermore, the FMT-acquired obese phenotype could be reverted by cohousing with lean rodents as a result of invasion by specific strains of Bacteroidetes, but only in mice fed a low-fat diet, clearly illustrating the interplay between diet and microbiota[65]. The potential obesogenic gut microbiota is also evident in humans, with reports of new-onset obesity in patients with recurrent Clostridium difficile after FMT from overweight donors[66].
Gut microbiota has a paramount role in the regulation of whole-body energy homeostasis and may induce obesity through direct host interactions or indirectly via microbial metabolites.
The gut microbiota is able to ferment indigestible carbohydrates into SCFA, which accounts for roughly 10% of harvested energy from diet[37]. Acetate is the main source of energy from SCFA and the most obesogenic one, being able to induce hyperphagia (through induction of ghrelin and glucose-stimulated insulin secretion), hypertriglyceridemia, IR, hepatic and adipocyte lipogenesis, and ectopic lipid deposition in the liver and skeletal muscle[67,68]. Conversely, butyrate and propionate are anti-obesogenic, both increasing the expression of the anorexigenic adipokine leptin[69,70] and gut hormones peptide tyrosine-tyrosine (PYY) and glucagon-like peptide-1 (GLP-1)[71]. Butyrate enhances energy expenditure by promoting mitochondrial activity and upregulating genes for lipolysis and fatty acid oxidation[72]. Propionate directly induces intestinal gluconeogenesis, which abrogates adiposity and weight gain, independently of food intake, through the activation of gut–brain neural circuits mediated by portal vein glucose sensors[73]. Butyrate is mainly produced by the Firmicutes Faecalibacterium prausnitzii, Eubacterium rectale, and Rosuberia intestinalis. Acetate is also produced by Bacteroidetes such as Bacteroides and Prevotella, as well as Lactobacillus, Bifidobacterium, and Akkermansia[70,74].
Gut microbiota downregulates the intestinal expression of fasting-induced adipose factor (FIAF). FIAF is a lipoprotein lipase inhibitor that inhibits adipose tissue and hepatic cellular uptake of fatty acids from circulating lipoproteins. As such, FIAF inhibition by the gut microbiota promotes liver steatogenesis and expansion of the adipose tissue[61].
The interplay between dietary tryptophan and gut microbiota may also contribute to obesity. In the gut, the main metabolic pathway of tryptophan is the kynurenine pathway, which results in different bioactive metabolites. Several gut bacteria have the potential to shunt tryptophan into two minor pathways: the aryl hydrocarbon receptor (AhR) pathway directly by metabolizing it into indoles (e.g., Lactobacillus) and the serotonin pathway indirectly by SCFA and secondary bile acids regulation of serotonin synthesis (e.g., Prevotella)[75,76]. The AhR pathway culminates in the production of GLP-1 and IL-22, the latter being able to decrease mucosal inflammation and permeability by inducing resistance to Candida albicans colonization[77,78]. Serotonin also acts on energy homeostasis by acting in brain reward centers and increasing lipolysis in white adipose tissue while decreasing thermogenesis in brown adipose tissue[79,80].
Dysbiota may also modulate the effects of the gut microbiota on bile acids, which may impact metabolism and energy homeostasis[81,82]. For example, an impairment in the production of secondary bile acids leads to the accumulation of primary bile acids with proinflammatory and leaky gut-inducing effects[83].
The gut microbiota is a main regulator of the intestinal mucosal barrier integrity, and gut dysbiota can induce a leaky gut through direct mucosal inflammation or indirectly via microbe metabolites[84]. The increased permeability to microbial products such as lipopolysaccharides (LPS) has been dubbed metabolic endotoxemia, which contributes to the inflammatory state that characterizes obesity, but it also has profound metabolic effects, inducing IR, leptin resistance, adipose tissue expansion, and hepatic steatosis[85-87]. Furthermore, gut inflammation increases its vascularization and enhances nutrient absorption[88].
Recent research is gathering information to characterize obesity-associated gut microbiota. Obesity is characterized by decreased gut microbiota richness and diversity[89], with a corresponding decrease in microbiome gene count[90].
Obesity-associated gut microbiota seems not only quantitatively but also qualitatively different. Most, but not all, studies have described an increased Firmicutes/Bacteroidetes ratio[91,92]. The huge variability between studies suggests that more than a taxonomic core, there is a microbiome core for obesity that is associated with higher efficiency in energy harvesting from diet and proinflammatory states[89,93,94]. However, some taxonomic associations have been fairly consistent, with obesity being associated with decreased relative abundance in the families Rikenellaceae and Christensenellaceae and the genera Bifidobacterium, Oscillospira, Faecalibacterium, and Akkermansia, all of which are involved in SCFA metabolism, mucosal integrity, and protection against a leaky gut[70]. Conversely, obesity has been associated with increased relative abundance in the families Prevotellaceae and Coriobacteriacea and the genera Roseburia and Eubacterium. H2-oxidizing methanogenic Archaea with the potential for higher carbohydrate fermentation efficiency also seem to be increased[70,94,95], as well as pathogenic proinflammatory Gram-negative bacteria, such as Escherichia or Shigella[94].
The diet seems to have a major role in the acquisition of an obese-associated gut microbiota[96], with high-fat, high-sucrose Western diets promoting a shifting microbiota composition, depleting “favorable” bacteria (e.g., Bifidobacterium and Faecalibacterium) while replenishing “unfavorable” bacteria (e.g., Blautia and Acinetobacter)[97].
GUT MICROBIOTA AND DIABETES MELLITUS
Type 2 diabetes mellitus (T2DM) shares behavioral risk factors with obesity, as well as similar dysbiota profiles.
Human observational studies have shown a lower - but not -diversity in T2DM patients[98,99]. Furthermore, a recent metaanalysis showed that T2DM is associated with an increased abundance of Firmicutes (e.g., Veillonellacea) and decreased Bacterioidetes[98]. At the genus level, the most striking associations were an increased relative abundance of Ruminococcus, Coprococcus, Eubacteria, Blautia, and Prevotella, whereas potentially protective bacteria were Dorea, Bifidobacterium, Bacteroides, Faecalibacterium, Akkermansia, and Roseburia[98-100].
Collectively, T2DM patients have a decrease in butyrate-producing bacteria and increased expression of microbiota genes related to inflammation and oxidative stress, leading to a leaky gut[101,102].
Increased intestinal permeability leading to endotoxemia and a continuous state of low-grade inflammation can induce IR[103]. For example, LPS binding to TLR4 induces IR by direct phosphorylation of insulin receptor[85,104,105]. Indeed, Roseburia, Bacteroides, and Akkermansia, which seem to be depleted in T2DM, either induce anti-inflammatory or inhibit proinflammatory cytokines[28,106,107]. Bacteroides and Akkermansia muciniphila further protect from the leaky gut by directly strengthening epithelial tight junctions[29,108].
Gut dysbiosis may also have a profound effect on glucose metabolism and overall energy homeostasis[100]. For example, the potentially anti-diabetogenic Bifidobacterium induces gut secretion of GLP-1 and PYY, promoting insulin sensitivity and beta cell function[109,110]. Microbial metabolites also modulate glucose homeostasis. For example, regarding SCFA, propionate reduces insulin sensitivity, leading to compensatory hyperinsulinemia[111]; acetate increases glucose-stimulated insulin secretion and weight gain[67]; and butyrate has insulin-sensitizing actions by indirectly promoting the gut barrier abrogating metabolic endotoxemia[112] and through the induction of insulin-sensitizing gut hormones GLP-1, PYY, and GLP-2[113].
Prevotella copri is associated with increased risk for T2DM, and the link may be the production of BCAAs[114], which are able to induce mTORC1-dependent IR[115].
Trimethylamine-N-oxide (TMAO) may be another link between gut dysbiota and T2DM[116]. TMAO induces IR through ER stress-induced FoxO1 expression[117,118]. Dietary choline can be used for phosphatidylcholine synthesis, oxidized into betaine or metabolized into trimethylamine (TMA) by gut microbiota, mainly Firmicutes and Proteobacteria such as Escherichia coli[119]. TMA is further oxidized in the liver to TMAO by hepatic flavin-containing monooxygenases (FMAO3). TMAO has been consistently associated with T2DM[120] and an increased cardiovascular risk[121,122].
Lastly, T2DM patients have lower levels of secondary biliary acids, decreasing the contribution of the latter to insulin sensitivity via TGR5 induction of GLP-1 secretion[123].
Further supporting the role of gut dysbiosis in T2DM pathogenesis, a small study with FMT in obese patients improved insulin sensitivity and glucose tolerance after 12 weeks, independent of weight loss[124], even though this was not consistently replicated by others[125-127].
GUT MICROBIOTA AND MAFLD
MAFLD is the hepatic manifestation of metabolic syndrome with strong associations with obesity and T2DM[4]. Unsurprisingly, it shares common core pathogenesis, which extends to sharing the gut microbiota[5]. However, not all MAFLD patients are obese or overweight[128], and one third of T2DM patients will not present MAFLD[129]. Predictably, specific microbiota features may be associated with MAFLD beyond the associations shared with metabolic dysfunction.
There is strong indirect evidence that MAFLD is associated with gut dysbiosis. Similar to obesity, MAFLD can be a transmissible disease in rodents by cohousing or direct FMT of microbiota-devoid mice from mice with genetically or diet-induced hepatic steatosis or from patients with nonalcoholic steatohepatitis (NASH), even when those mice were fed a healthy isocaloric diet[61,130-134]. Furthermore, antibiotic treatment prevents the development of hepatic steatosis in dietary and genetically induced obesity in animal models[135,136], as well as in obese patients submitted to intestinal bypass bariatric surgery[137]. There is also strong preclinical and clinical evidence of an association among small intestine bacterial overgrowth[138,139], leaky gut[140], and MAFLD. Although only up to 40% of patients with MAFLD present an increase in intestinal permeability, this is six folds higher than the level expected in healthy subjects[141-143].
Studies comparing gut microbiota from healthy subjects, obese patients, and patients with MAFLD found greater differences between obese and non-obese phenotypes and greater similarities between obese and MAFLD phenotypes[144]. Both obesity and MAFLD seem to be associated with decreased bacterial species diversity[145], although the diversity is even lower in patients with MAFLD[146]. In obesity, at the phylum level, an increased Firmicutes/Bacteroidetes ratio has been consistently described. In MAFLD, the relative abundance of Firmicutes and Bacteroidetes was variable[144,147-150]; however, when direct comparisons of the microbiota between obese patients and MAFLD patients were made, their ratio was similar in some studies[144,146] or reversed with decreased Firmicutes and increased Bacteroidetes in MAFLD patients, particularly in patients with steatohepatitis[134,151,152]. At the family level, both obesity and MAFLD are associated with a relative decrease in Lachnospiraceae and Ruminocococcaceae[153], and, at the genus level, there was a decrease in Faecalibacterium as well as an increase in Prevotella in some but not all cohorts[6,143,152,154].
Faecalibacterium, Eubacterium, Rosuberia, and Coprococcus are all butyrate-producing bacteria that consistently were shown to be decreased in MAFLD[147,152]. Faecalibacterium praunitzii, in particular, belongs to the Ruminococcaceae family[155], accounts for over 5% of the total gut microbiota in healthy humans[156], and regulates hepatic fat content, upregulating fatty acids oxidation and increasing hepatic sensitivity to adiponectin[157]. It also secretes anti-inflammatory compounds such as salicylic acid[158] and microbial anti-inflammatory molecule (MAM) protein[159], which abrogate adipose tissue inflammation[155].
Akkermansia depletion is associated with disruption of gut epithelial barrier, is characteristic of obesity and T2DM, and was also reported in MAFLD[147].
Similar to T2DM and the metabolic syndrome, TMAO levels increase (with correspondent decreases in betaine) in parallel with the severity of hepatic steatosis[160]. Prevotella gut colonization seems to be associated with increased levels of TMAO[161], and several studies described increased Prevotella abundance in MAFLD patients[143]. The increase in TMAO in MAFLD may result in metabolic dysfunction-independent steatogenic effects, since TMAO synthesis shunts choline away from the synthesis of betaine (a methyl donor with antioxidant and anti-inflammatory properties[162]) and phosphatidylcholine (necessary for lipoprotein export from the liver[163]). Furthermore, TMAO has direct steatogenic effects in the liver by suppressing bile acid-mediated hepatic FXR signaling, which results in SREPBP-1c-mediated induction of lipogenesis. This was matched with a shift in the composition of the bile acid pool, with a decrease in potent FXR agonists tauroCDCA and glycolithocholic acids and an increase in FXR antagonist taurocholic acid[164]. Different bacteria belonging to Firmicutes and Proteobacteria are able to produce TMA, such as some Clostridium species, Escherichia fergusonii, and Proteus penneri[165]. Some of those bacteria have been associated with MAFLD. For example, Escherichia fergusonii was associated with non-obese hepatic steatosis in animal models and humans[166].
Gut dysbiota may have steatogenic effects independent of the ones leading to obesity and IR/T2DM, mainly through bile acid metabolism and microbiota end-products/metabolites. Indeed, when comparing morbid non-diabetic obese women with and without steatosis, a 75% shared variation between the gut microbiome and the molecular phenomics was found (i.e., the sum of hepatic transcriptome and plasma and urine metabolome), which was associated with steatosis, suggesting a causal effect[134].
Some bacteria produce ethanol, which is directly steatogenic[144]. Accordingly, several studies, including on pediatric populations, showed MAFLD/NASH to be associated with increased endogenous ethanol levels[144,167]. Examples of ethanol-producing bacteria are Proteobacteria such as Escherichia coli[168]. Interestingly, an increase in Proteobacteria relative abundance is one of the most consistent findings in MAFLD patients[6,151,153,154]. An increase in Proteobacteria, and particularly Escherichia coli, is a distinguishing gut microbiota trait between obese and MAFLD/NASH patients[143,144,146,147,169]. Of note, an overgrowth of Escherichia coli in the gut microbiota has been linked to subtherapeutic doses of antibiotics that may be a contaminant of Western diets[170]. Additionally, in a Chinese cohort of MAFLD patients, 60% presented an increased relative abundance of another Proteobacteria: high alcohol-producing Klebsiella pneumoniae[171]. From that cohort, one patient presented severe NASH and auto-brewing syndrome; that is, he developed ultra-high alcohol blood concentration after an alcohol-free, high-carbohydrate diet[172]. Transplantation of isolates of high alcohol producing Klebsiella pneumonia from that patient into a rodent recipient induced hepatic steatosis[171,173]. An increased relative abundance of Klebsiella was corroborated by other groups[147].
Proteobacteria are also relevant as a source of LPS[134]. Indeed, the microbiome of patients with MAFLD/steatohepatitis is characteristically enriched in genes for LPS synthesis[151].
T2DM is associated with decreased Lactobacillus[174]; however, in MAFLD, different groups described an actual increase in Lactobacillus relative abundance[143,151,153]. The increased abundance of Lactobacillus in MAFLD is counterintuitive since it is a known probiotic that competitively inhibits pathogens, heightens the epithelial barrier function, and has immunomodulatory properties[175]. However, Lactobacillus also produces volatile organ compounds and ethanol, which are known potentiators of steatogenesis and steatohepatitis[176]. Indeed, the Lactobacillus genus comprises more than 180 species with different sugar fermentation properties, the end result of some species being predominantly lactic acid (L. acidophilus and L. salivarius) and others ethanol (L. casei, L. brevis, and L. plantarum)[5].
Gut microbiota can modulate the development of MAFLD through other microbial byproducts. MAFLD-associated microbiota also showed an increased genetic potential for hepatic inflammation and deregulation of AAA (tryptophan, tyrosine, and phenylalanine) and BCAA (valine, leucin, and isoleucine)[134]. BCAAs are associated with obesity and IR[114]. Steatohepatitis has been associated with an increase in the genus Bacteroides and Prevotella copri which have the ability to produce BCAA[143,154,177]. Another microbiota-associated metabolite that was strongly associated with MAFLD is the AAA-derived phenylacetic acid, which showed direct steatogenic effects on hepatocytes in vitro, promoting hepatic lipids uptake (by induction of lipoprotein lipase), de novo lipogenesis (by inducing fatty acids synthase), and inhibiting the insulin receptor pathway[134]. 2-butanone seems to decrease in NAFLD[153] but increase in steatohepatitis, which may be the result of increased Streptococcus pneumonia[143,146,154]. 2-butanone is a strong inductor of CYP, and even though it is not hepatotoxic per se, it potentiates hepatic lesions from toxic-associated steatohepatitis[178,179].
The effects of the gut microbiota on bile acid metabolism also have a crucial role in the development of MAFLD. MAFLD is associated with an increased abundance of bacteria that produce bile salt hydrolases and hence deconjugate bile acids (e.g., Clostridium, Lactobacillus, Escherichia, and Bacteroides), as well as bacteria that facilitate the conversion of primary to secondary bile acids (predominantly Clostridium species[143,148,180]). Primary bile acids activate, whereas secondary bile acids inhibit FXR[142]. FXR activity is known to be decreased in MAFLD, which is associated with an increased synthesis of hepatic bile acids, metabolic deregulation, steatogenesis, inflammation, and possibly fibrogenesis in the liver[181].
Non-obese MAFLD shares microbiota signatures with obese MAFLD. In fact, some studies even found an obese-associated microbiota in lean patients with MAFLD, such as an increased Firmicutes/Bacterioidetes ratio[182] and a decrease in Lachnospiraceae (e.g., Coprococcus) and Ruminococcaceae (e.g., Ruminococcus). However, other microbiota associations with lean MAFLD suggest a different microbiota profile that could induce steatogenesis by mechanisms unrelated to the metabolic syndrome[155]. For example, lean versus obese NAFLD is associated with decreased Lactobacillus abundance[182,183]. Furthermore, non-obese steatohepatitis is associated with a different mycobiome, with a lower fungal richness, increased abundance of Candida albicans and Mucor spp, and decreased Saccharomyces cerevisiae[184]. C. albicans is known to induce a Th17 proinflammatory response[185], and its richness is associated more with hepatic inflammation than steatosis[184].
GUT MICROBIOTA AND PROGRESSION OF MAFLD
The presence and severity of liver fibrosis dictate the risk for progression to liver cirrhosis and morbi-mortality[186]. The gut microbiota changes according to the severity of liver disease, from mild to significant fibrosis, compensated cirrhosis, and decompensated cirrhosis. The differences found in the gut microbiota allowed the proposal of gut microbiome-based metagenomic signatures for MAFLD with advanced fibrosis and cirrhosis[169,187,188], with apparent great diagnostic accuracy.
Interestingly, while - and -diversity seem to decrease with mild disease, they paradoxically increase with severe disease, with increased dispersion in cirrhosis[169].
Advanced fibrosis, as compared to absence or mild fibrosis, has been associated with increased anaerobic bacteria (e.g., Bacteroides vulgatus, Holdemanella, and Prevotella copri, even though an increase in the latter was not consensual[22,177]) and Gram-negative bacteria (e.g., the Proteobacteria Escherichia coli and
Several reports showed an association between advanced fibrosis and decreased Ruminococcus obeum[22,155,177] and Eubacterium rectale[187]. Both species are fiber-fermenting bacteria with the ability to produce SCFA and have also been negatively associated with the development of primary sclerosing cholangitis in patients with inflammatory bowel disease[195]. Indeed, SCFAs inhibit hepatic inflammation (a major driver of fibrogenesis) via promoting intestinal barrier integrity and directly with anti-inflammatory actions on Kupffer cells[196].
The metagenome associated with advanced fibrosis is correlated with bacterial changes. For example, advanced fibrosis was associated with the anaerobic bacteria derived 3-phenylpropanoate[187]. Increased abundance of the anaerobic bacteria Prevotella and Holdemanella is also associated with oxidative stress and a proinflammatory environment, as a consequence of changes in metabolic pathways such as urea cycle and vitamin B biosynthesis[190,197].
Besides bacteria, the virome in patients with advanced fibrosis is different, with a decreased variability and a decreased relative abundance of bacteriophages, which may have a direct impact on modulating the taxonomic composition of the bacterial microbiota[198]. In addition, regarding mycobiome, Candida albicans relative abundance and immunogenicity seem to increase in patients with advanced fibrosis[184].
Cirrhosis seems to be associated with invasion of the gut by oral commensals, such as Prevotella, Veillonella, and Streptococcus, possibly through altered bile acid metabolism[199,200]. Conversely, there is a further decrease in beneficial commensals such as F. prausnitzii and Coprococcus[188,199].
Decompensated cirrhosis is associated with a further dramatic shift in gut microbiota, with an increased abundance of pathogenic Enterococcaceae and Enterobacteriaceae and decreased Bacteroidaceae and the Lachnospiracea Clostridiales IV[201,202], which is correlated with endotoxemia[203]. Again, when cirrhosis decompensates, there is a decrease in the synthesis of bile acids and their fecal concentrations, which abrogates bile acids’ negative effect on Gram-negative bacteria, increasing the ratio of Gram-negative to Gram-positive bacteria (e.g., Lachnospiraceae)[149].
MAFLD-associated hepatocellular carcinoma is associated with intestinal inflammation, as evidenced by increased fecal calprotectin and increased cytokines IL-8, IL-13, and CCL-3/4/5, which is correlated with a decrease in Akkermansia and Bifidobacterium[200]. Furthermore, there is also an increase in pathogenic Enterobactereacea compared to MAFLD cirrhosis without hepatocellular carcinoma. In contrast to the increased intestinal inflammation, MAFLD-HCC microbiota induces a peripheral T cell immunosuppressive phenotype, expansion of regulatory T cells, and attenuation of effector CD8+ T cells, which may be responsible for a decreased tumoral immunosurveillance[204].
MAFLD-associated hepatocellular carcinoma is also associated with increased total bile acids and serum FGF-19, which is correlated with an increase in Lactobacilli[205]. A gut dysbiota-dependent increase in DCA has been described in MAFLD cirrhosis and may have pro-tumoral effects. Indeed, DCA induces a senescence-associated secretory phenotype in hepatic stellate cells, which elicits the secretion of proinflammatory and tumor-promoting factors[206]. Furthermore, DCA further inhibits tumoral immunosurveillance by inhibiting the expression of CXCL16 in hepatic sinusoidal cells, hence recruiting fewer anti-tumoral CRCR6+ NKT cells[207] [Figure 1].
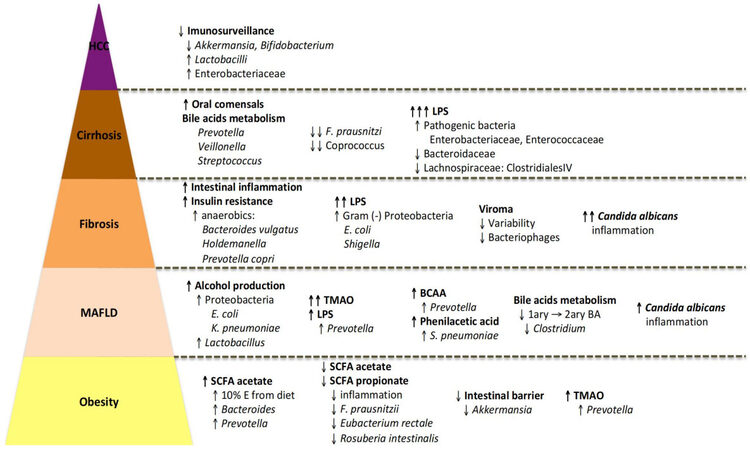
Figure 1. Step-by-step acquisition of unhealthy patterns of gut dysbiosis with the severity of liver disease in patients with MAFLD. TMAO: Trimethylamine-N-oxide; MAFLD: metabolic-associated fatty liver disease; HCC: hepatocellular carcinoma; LPS: lipopolysaccharides; SCFA: short-chain fatty acids.
INTERVENTION IN GUT MICROBIOTA FOR THE TREATMENT OF MAFLD
As of today, no treatment has been approved for MAFLD, and gut dysbiosis is a highly appealing treatment target. Microbiome-targeted therapies include different strategies, such as probiotics and prebiotics, antibiotics, FMT, and bacteriophages. Furthermore, treatment strategies applied today, for example, diet and exercise, bariatric surgery, and some anti-diabetic agents such as metformin[208] and liraglutide[209], act through modulation of the microbiota. Metformin is able to alter the gut microbiota composition in a way that leads to an improvement in glucose intolerance[208]. Indeed, metformin has a dose-dependent antimicrobial effect with a decrease of specific bacteria[210] such as Bacterioides fragilis, and depletion of Bacterioides fragilis is associated with inhibited FXR signaling mediated by glycoursodeoxycholic acid[211]. Metformin also promotes the growth of specific bacteria such as Akkermansia muciniphila, which is known to have beneficial metabolic and anti-inflammatory effects[212]. There is also scarce preclinical evidence of treatment of Chinese herbs such as Jiangzi granules and Ixeris chinensis showing beneficial effects on MAFLD through the modulation of the gut microbiota[213-215]. Furthermore, alcohol consumption promotes Prevotella growth, high protein intake decreases Bacteroides abundance[216], and exercise increases SCFA-producing bacteria and anti-inflammatory F. prausnitzii and A. muciniphila[217].
Probiotics are live non-pathogenic microorganisms that induce health benefits in humans when in appropriate quantities. The most used probiotics are Lactobacilli and Bifidobacteria[218]. Prebiotics are fermentable dietary fibers (such as inulin and fructo-oligosaccharides) that selectively promote the growth of probiotics. The combination of probiotics and prebiotics is dubbed synbiotics[8]. Small short-term clinical trials have evaluated the effect of probiotics on MAFLD. Different metaanalyses showed probiotic therapy to result in biochemical improvement of aminotransferases and lipid profile, as well as of steatosis and fibrosis by non-invasive techniques, while there are no data on liver histology[218-220]. Comparing different studies with probiotics is hampered by the fact that each study uses different formulations of probiotics. Furthermore, even though we can find some patterns of dysbiosis in MAFLD, the gut microbiota is unique for each patient, which anticipates a highly variable effect of probiotics. The effect of probiotics is also expected to be only transient, since there is no evidence of persistent probiotic engraftment with a persistent shift in the microbiota composition[221]. The best approach would be personalized probiotics therapy, with the selection of specific commensal bacteria that would already be present in the individual baseline microbiota to allow better engraftment[222].
The evidence for prebiotics in the treatment of MAFLD is even scarcer, with very small trials in adults suggesting a mild effect on body mass index and aminotransferases levels[223].
Less is known regarding the effect of antibiotics on MAFLD. Small studies administered rifaximin, a gut-specific antimicrobial agent, to patients with NASH and showed a mild reduction in body mass index, endotoxemia, and liver enzymes at doses of 1200 mg per day[224,225], while there was no effect for lower doses[226].
FMT consists of collecting stools from a healthy donor and transferring it to a patient. The transference can be accomplished by colonoscopy, nasogastric tube, enema, or gastric-resistant pills[42]. Small studies already tested the effect of FMT on the metabolic syndrome and showed benefits in glucose and lipid metabolism, albeit with no effect on body weight[126,127,227,228]. Regarding MAFLD, a small pilot study performed FMT in 21 patients and did not achieve improvements in IR or liver fat content, but a reduction in small intestine permeability was achieved, suggesting a beneficial effect on the intestinal epithelial barrier[229].
An innovative way to modulate microbiota is through bacteriophages therapy. Bacteriophages are ubiquitous viruses that can infect and destroy bacteria, often with species-level specificity. This approach has not been studied in MAFLD[230]. However, preclinical data on alcoholic hepatitis approached with a bacteriophage that specifically targets cytolytic Enterococcus faecalis show promising results[231].
CONCLUSIONS
Dramatizing, we are only 50% humans, since roughly as many bacteria as the human cells that compose our body colonize us. Half of the microbes have profound effects on our health and disease pathogenesis. This is the case for the development of the metabolic syndrome and its liver associated disease, MAFLD. Furthermore, gut dysbiosis has the ability to promote MAFLD by direct steatogenic metabolic syndrome-independent effects.
The study of gut microbiota has benefited from the tremendous recent advances in molecular testing, and today we can fairly accurately characterize an individual’s microbiota and microbiome. However, generalizations for a population and even more so for different populations are hampered by inter- and intra-individual differences, as well as geographic differences. As such, even though microbiota-targeted therapies for MAFLD are quite appealing, they are still far from clinical practice application. The field is moving towards a personalized microbiome targeted therapy, in which first we would characterize the individual gut microbiota, and afterwards, we would modulate the microbiota in a species-specific fashion in order to modulate the individual gut microbiota into a healthier, metabolically beneficial, and anti-steatogenic flora.
DECLARATIONS
Authors’ contributionsConceptualized the manuscript: Machado MV
Wrote the manuscript: Machado MV, Sousa P
Availability of data and materialsNot applicable.
Financial support and sponsorshipNot applicable.
Conflicts of interestAll authors declared that there are no conflicts of interest.
Ethical approval and consent to participateNot applicable.
Consent for publicationNot applicable.
Copyright© The Author(s) 2022
REFERENCES
1. Le MH, Yeo YH, Li X, et al. 2019 Global NAFLD Prevalence: a systematic review and Meta-analysis. Clin Gastroenterol Hepatol ;2021:S1542-3565(21)01280.
2. Younossi ZM, Stepanova M, Ong J, et al. Nonalcoholic steatohepatitis is the most rapidly increasing indication for liver transplantation in the United States. Clin Gastroenterol Hepatol 2021;19:580-589.e5.
3. Eslam M, Newsome PN, Sarin SK, et al. A new definition for metabolic dysfunction-associated fatty liver disease: an international expert consensus statement. J Hepatol 2020;73:202-9.
4. Machado MV, Diehl AM. Pathogenesis of nonalcoholic steatohepatitis. Gastroenterology 2016;150:1769-77.
5. Machado MV, Cortez-Pinto H. Diet, microbiota, obesity, and NAFLD: a dangerous quartet. Int J Mol Sci 2016;17:481.
6. Aron-Wisnewsky J, Vigliotti C, Witjes J, et al. Gut microbiota and human NAFLD: disentangling microbial signatures from metabolic disorders. Nat Rev Gastroenterol Hepatol 2020;17:279-97.
7. Fraher MH, O'Toole PW, Quigley EM. Techniques used to characterize the gut microbiota: a guide for the clinician. Nat Rev Gastroenterol Hepatol 2012;9:312-22.
8. Lynch SV, Pedersen O. The human intestinal microbiome in health and disease. N Engl J Med 2016;375:2369-79.
9. Langille MG, Zaneveld J, Caporaso JG, et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat Biotechnol 2013;31:814-21.
10. Gilbert JA, Blaser MJ, Caporaso JG, Jansson JK, Lynch SV, Knight R. Current understanding of the human microbiome. Nat Med 2018;24:392-400.
11. Knight R, Vrbanac A, Taylor BC, et al. Best practices for analysing microbiomes. Nat Rev Microbiol 2018;16:410-22.
12. Sharpton SR, Ajmera V, Loomba R. Emerging role of the gut microbiome in nonalcoholic fatty liver disease: from composition to function. Clin Gastroenterol Hepatol 2019;17:296-306.
13. Franzosa EA, Huang K, Meadow JF, et al. Identifying personal microbiomes using metagenomic codes. Proc Natl Acad Sci U S A 2015;112:E2930-8.
14. Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature 2012;489:220-30.
15. Arumugam M, Raes J, Pelletier E, et al. MetaHIT Consortium (additional members). Enterotypes of the human gut microbiome. Nature 2011;473:174-80.
16. Ding RX, Goh WR, Wu RN, et al. Revisit gut microbiota and its impact on human health and disease. J Food Drug Anal 2019;27:623-31.
17. Rodriguez-Valera F, Martin-Cuadrado AB, Rodriguez-Brito B, et al. Explaining microbial population genomics through phage predation. Nat Rev Microbiol 2009;7:828-36.
18. Quigley EM. Gut microbiota and the role of probiotics in therapy. Curr Opin Pharmacol 2011;11:593-603.
19. Gill SR, Pop M, Deboy RT, et al. Metagenomic analysis of the human distal gut microbiome. Science 2006;312:1355-9.
20. Hooper LV, Gordon JI. Commensal host-bacterial relationships in the gut. Science 2001;292:1115-8.
21. Qin J, Li R, Raes J, et al. MetaHIT Consortium. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010;464:59-65.
22. Smirnova E, Muthiah MD, Narayan N, et al. Metabolic reprogramming of the intestinal microbiome with functional bile acid changes underlie the development of NAFLD. Hepatology 2022; doi: 10.1002/hep.32568.
23. Ramirez J, Guarner F, Bustos Fernandez L, Maruy A, Sdepanian VL, Cohen H. Antibiotics as major disruptors of gut microbiota. Front Cell Infect Microbiol 2020;10:572912.
24. Chen Y, Zhou J, Wang L. Role and mechanism of gut microbiota in human disease. Front Cell Infect Microbiol 2021;11:625913.
25. Shreiner AB, Kao JY, Young VB. The gut microbiome in health and in disease. Curr Opin Gastroenterol 2015;31:69-75.
26. Walters KE, Martiny JBH. Alpha-, beta-, and gamma-diversity of bacteria varies across habitats. PLoS One 2020;15:e0233872.
27. Allam-Ndoul B, Castonguay-Paradis S, Veilleux A. Gut microbiota and intestinal trans-epithelial permeability. Int J Mol Sci 2020;21:6402.
28. Plovier H, Everard A, Druart C, et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat Med 2017;23:107-13.
29. Chelakkot C, Choi Y, Kim DK, et al. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp Mol Med 2018;50:e450.
30. McFarland LV. Antibiotic-associated diarrhea: epidemiology, trends and treatment. Future Microbiol 2008;3:563-78.
31. Khosravi A, Yáñez A, Price JG, et al. Gut microbiota promote hematopoiesis to control bacterial infection. Cell Host Microbe 2014;15:374-81.
32. Jandhyala SM, Talukdar R, Subramanyam C, Vuyyuru H, Sasikala M, Nageshwar Reddy D. Role of the normal gut microbiota. World J Gastroenterol 2015;21:8787-803.
33. Macfarlane S, Macfarlane GT. Regulation of short-chain fatty acid production. Proc Nutr Soc 2003;62:67-72.
34. Wen L, Wong FS. Dietary short-chain fatty acids protect against type 1 diabetes. Nat Immunol 2017;18:484-6.
35. Chambers ES, Morrison DJ, Frost G. Control of appetite and energy intake by SCFA: what are the potential underlying mechanisms? Proc Nutr Soc 2015;74:328-36.
36. Hu J, Lin S, Zheng B, Cheung PCK. Short-chain fatty acids in control of energy metabolism. Crit Rev Food Sci Nutr 2018;58:1243-9.
37. Brahe LK, Astrup A, Larsen LH. Is butyrate the link between diet, intestinal microbiota and obesity-related metabolic diseases? Obes Rev 2013;14:950-9.
38. Wang HB, Wang PY, Wang X, Wan YL, Liu YC. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein Claudin-1 transcription. Dig Dis Sci 2012;57:3126-35.
39. Rowland I, Gibson G, Heinken A, et al. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr 2018;57:1-24.
40. Hill MJ. Intestinal flora and endogenous vitamin synthesis. Eur J Cancer Prev 1997;6 Suppl 1:S43-5.
41. Macfarlane GT, Cummings JH, Allison C. Protein degradation by human intestinal bacteria. J Gen Microbiol 1986;132:1647-56.
42. Cani PD, Van Hul M, Lefort C, Depommier C, Rastelli M, Everard A. Microbial regulation of organismal energy homeostasis. Nat Metab 2019;1:34-46.
43. Begley M, Gahan CG, Hill C. The interaction between bacteria and bile. FEMS Microbiol Rev 2005;29:625-51.
44. García-Cañaveras JC, Donato MT, Castell JV, Lahoz A. Targeted profiling of circulating and hepatic bile acids in human, mouse, and rat using a UPLC-MRM-MS-validated method. J Lipid Res 2012;53:2231-41.
45. Ahmad TR, Haeusler RA. Bile acids in glucose metabolism and insulin signalling - mechanisms and research needs. Nat Rev Endocrinol 2019;15:701-12.
46. Ðanić M, Stanimirov B, Pavlović N, et al. Pharmacological applications of bile acids and their derivatives in the treatment of metabolic syndrome. Front Pharmacol 2018;9:1382.
47. Donaldson GP, Lee SM, Mazmanian SK. Gut biogeography of the bacterial microbiota. Nat Rev Microbiol 2016;14:20-32.
48. Wu GD, Chen J, Hoffmann C, et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011;334:105-8.
49. Allaband C, McDonald D, Vázquez-Baeza Y, et al. Microbiome 101: studying, analyzing, and interpreting gut microbiome data for clinicians. Clin Gastroenterol Hepatol 2019;17:218-30.
50. Dominguez-Bello MG, Costello EK, Contreras M, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A 2010;107:11971-5.
51. Mackie RI, Sghir A, Gaskins HR. Developmental microbial ecology of the neonatal gastrointestinal tract. Am J Clin Nutr 1999;69:1035S-45S.
52. Hildebrandt MA, Hoffmann C, Sherrill-Mix SA, et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology 2009;137:1716-24.e1.
53. De Filippo C, Cavalieri D, Di Paola M, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A 2010;107:14691-6.
54. Hermanson JB, Fei N, Miyoshi S, et al. Dietary cholesterol-induced gut microbes drive nonalcoholic fatty liver disease pathogenesis in a murine model. The FASEB Journal 2022;36:fasebj.2022.36.S1.0R748.
55. He Y, Wu W, Zheng HM, et al. Regional variation limits applications of healthy gut microbiome reference ranges and disease models. Nat Med 2018;24:1532-5.
57. Sonnenburg JL, Sonnenburg ED. Vulnerability of the industrialized microbiota. Science 2019;366:eaaw9255.
58. Lim MY, You HJ, Yoon HS, et al. The effect of heritability and host genetics on the gut microbiota and metabolic syndrome. Gut 2017;66:1031-8.
59. Goodrich JK, Davenport ER, Beaumont M, et al. Genetic determinants of the gut microbiome in UK twins. Cell Host Microbe 2016;19:731-43.
60. Qin Y, Havulinna AS, Liu Y, et al. Combined effects of host genetics and diet on human gut microbiota and incident disease in a single population cohort. Nat Genet 2022;54:134-42.
61. Bäckhed F, Ding H, Wang T, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A 2004;101:15718-23.
62. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006;444:1027-31.
63. Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci U S A 2007;104:979-84.
64. Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, Gordon JI. The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci Transl Med 2009;1:6ra14.
65. Ridaura VK, Faith JJ, Rey FE, et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013;341:1241214.
66. Alang N, Kelly CR. Weight gain after fecal microbiota transplantation. Open Forum Infect Dis 2015;2:ofv004.
67. Perry RJ, Peng L, Barry NA, et al. Acetate mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature 2016;534:213-7.
68. Chakraborti CK. New-found link between microbiota and obesity. World J Gastrointest Pathophysiol 2015;6:110-9.
69. Xiong Y, Miyamoto N, Shibata K, et al. Short-chain fatty acids stimulate leptin production in adipocytes through the G protein-coupled receptor GPR41. Proc Natl Acad Sci U S A 2004;101:1045-50.
70. Cunningham AL, Stephens JW, Harris DA. A review on gut microbiota: a central factor in the pathophysiology of obesity. Lipids Health Dis 2021;20:65.
71. Chambers ES, Viardot A, Psichas A, et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015;64:1744-54.
72. Coppola S, Avagliano C, Calignano A, Berni Canani R. The protective role of butyrate against obesity and obesity-related diseases. Molecules 2021;26:682.
73. De Vadder F, Kovatcheva-Datchary P, Goncalves D, et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014;156:84-96.
74. Louis P, Flint HJ. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol Lett 2009;294:1-8.
75. Taleb S. Tryptophan dietary impacts gut barrier and metabolic diseases. Front Immunol 2019;10:2113.
76. Agus A, Planchais J, Sokol H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe 2018;23:716-24.
77. Natividad JM, Agus A, Planchais J, et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab 2018;28:737-749.e4.
78. Zelante T, Iannitti RG, Cunha C, et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013;39:372-85.
79. Dong TS, Guan M, Mayer EA, et al. Obesity is associated with a distinct brain-gut microbiome signature that connects Prevotella and Bacteroides to the brain's reward center. Gut Microbes 2022;14:2051999.
80. Crane JD, Palanivel R, Mottillo EP, et al. Inhibiting peripheral serotonin synthesis reduces obesity and metabolic dysfunction by promoting brown adipose tissue thermogenesis. Nat Med 2015;21:166-72.
81. Li F, Jiang C, Krausz KW, et al. Microbiome remodelling leads to inhibition of intestinal farnesoid X receptor signalling and decreased obesity. Nat Commun 2013;4:2384.
82. Parséus A, Sommer N, Sommer F, et al. Microbiota-induced obesity requires farnesoid X receptor. Gut 2017;66:429-37.
83. Raimondi F, Santoro P, Barone MV, et al. Bile acids modulate tight junction structure and barrier function of Caco-2 monolayers via EGFR activation. Am J Physiol Gastrointest Liver Physiol 2008;294:G906-13.
84. Cani PD, Possemiers S, Van de Wiele T, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009;58:1091-103.
85. Cani PD, Amar J, Iglesias MA, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007;56:1761-72.
86. Pendyala S, Walker JM, Holt PR. A high-fat diet is associated with endotoxemia that originates from the gut. Gastroenterology 2012;142:1100-1101.e2.
87. Lartigue G, de la Serre CB, Espero E, Lee J, Raybould HE. Diet-induced obesity leads to the development of leptin resistance in vagal afferent neurons. Am J Physiol Endocrinol Metab 2011;301:E187-E95.
88. Ding S, Chi MM, Scull BP, et al. High-fat diet: bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. PLoS One 2010;5:e12191.
89. Turnbaugh PJ, Hamady M, Yatsunenko T, et al. A core gut microbiome in obese and lean twins. Nature 2009;457:480-4.
90. Chatelier E, Nielsen T, Qin J, et al; MetaHIT consortium. Richness of human gut microbiome correlates with metabolic markers. Nature 2013;500:541-6.
91. Walters WA, Xu Z, Knight R. Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett 2014;588:4223-33.
92. Pinart M, Dötsch A, Schlicht K, et al. Gut microbiome composition in obese and non-obese persons: a systematic review and meta-analysis. Nutrients 2021;14:12.
93. Vallianou N, Stratigou T, Christodoulatos GS, Dalamaga M. Understanding the role of the gut microbiome and microbial metabolites in obesity and obesity-associated metabolic disorders: current evidence and perspectives. Curr Obes Rep 2019;8:317-32.
94. Gao R, Zhu C, Li H, et al. Dysbiosis signatures of gut microbiota along the sequence from healthy, young patients to those with overweight and obesity. Obesity (Silver Spring) 2018;26:351-61.
95. Zhang H, DiBaise JK, Zuccolo A, et al. Human gut microbiota in obesity and after gastric bypass. Proc Natl Acad Sci U S A 2009;106:2365-70.
96. Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008;3:213-23.
97. Kong C, Gao R, Yan X, Huang L, Qin H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019;60:175-84.
98. Que Y, Cao M, He J, et al. Gut bacterial characteristics of patients with type 2 diabetes mellitus and the application potential. Front Immunol 2021;12:722206.
99. Wu H, Tremaroli V, Schmidt C, et al. The gut microbiota in prediabetes and diabetes: a population-based cross-sectional study. Cell Metab 2020;32:379-390.e3.
100. Gurung M, Li Z, You H, et al. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020;51:102590.
102. Palau-Rodriguez M, Tulipani S, Isabel Queipo-Ortuño M, Urpi-Sarda M, Tinahones FJ, Andres-Lacueva C. Metabolomic insights into the intricate gut microbial-host interaction in the development of obesity and type 2 diabetes. Front Microbiol 2015;6:1151.
103. Pussinen PJ, Havulinna AS, Lehto M, Sundvall J, Salomaa V. Endotoxemia is associated with an increased risk of incident diabetes. Diabetes Care 2011;34:392-7.
104. Shi H, Kokoeva MV, Inouye K, Tzameli I, Yin H, Flier JS. TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest 2006;116:3015-25.
106. Shen Z, Zhu C, Quan Y, et al. Insights into Roseburia intestinalis which alleviates experimental colitis pathology by inducing anti-inflammatory responses. J Gastroenterol Hepatol 2018;33:1751-60.
107. Chang YC, Ching YH, Chiu CC, et al. TLR2 and interleukin-10 are involved in Bacteroides fragilis-mediated prevention of DSS-induced colitis in gnotobiotic mice. PLoS One 2017;12:e0180025.
108. Yoshida N, Emoto T, Yamashita T, et al. Bacteroides vulgatus and bacteroides dorei reduce gut microbial lipopolysaccharide production and inhibit atherosclerosis. Circulation 2018;138:2486-98.
110. Kim SH, Huh CS, Choi ID, et al. The anti-diabetic activity of Bifidobacterium lactis HY8101 in vitro and in vivo. J Appl Microbiol 2014;117:834-45.
111. Tirosh A, Calay ES, Tuncman G, et al. The short-chain fatty acid propionate increases glucagon and FABP4 production, impairing insulin action in mice and humans. Sci Transl Med 2019;11:eaav0120.
112. Peng L, Li ZR, Green RS, Holzman IR, Lin J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J Nutr 2009;139:1619-25.
113. Arora T, Bäckhed F. The gut microbiota and metabolic disease: current understanding and future perspectives. J Intern Med 2016;280:339-49.
114. Pedersen HK, Gudmundsdottir V, Nielsen HB, et al. MetaHIT Consortium. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016;535:376-81.
115. Lynch CJ, Adams SH. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat Rev Endocrinol 2014;10:723-36.
116. Zhuang R, Ge X, Han L, et al. Gut microbe-generated metabolite trimethylamine N-oxide and the risk of diabetes: a systematic review and dose-response meta-analysis. Obes Rev 2019;20:883-94.
117. Chen S, Henderson A, Petriello MC, et al. Trimethylamine N-Oxide binds and activates PERK to promote metabolic dysfunction. Cell Metab 2019;30:1141-1151.e5.
118. Gao X, Liu X, Xu J, Xue C, Xue Y, Wang Y. Dietary trimethylamine N-oxide exacerbates impaired glucose tolerance in mice fed a high fat diet. J Biosci Bioeng 2014;118:476-81.
119. Arias N, Arboleya S, Allison J, et al. The Relationship between choline bioavailability from diet, intestinal microbiota composition, and its modulation of human diseases. Nutrients 2020;12:2340.
120. Dambrova M, Latkovskis G, Kuka J, et al. Diabetes is associated with higher trimethylamine N-oxide plasma levels. Exp Clin Endocrinol Diabetes 2016;124:251-6.
121. Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011;472:57-63.
122. Tang WH, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med 2013;368:1575-84.
123. Pathak P, Xie C, Nichols RG, et al. Intestine farnesoid X receptor agonist and the gut microbiota activate G-protein bile acid receptor-1 signaling to improve metabolism. Hepatology 2018;68:1574-88.
124. Allegretti JR, Kassam Z, Hurtado J, et al. Impact of fecal microbiota transplantation with capsules on the prevention of metabolic syndrome among patients with obesity. Hormones (Athens) 2021;20:209-11.
125. Zhang Z, Mocanu V, Cai C, et al. Impact of fecal microbiota transplantation on obesity and metabolic syndrome-a systematic review. Nutrients 2019;11:2291.
126. Yu EW, Gao L, Stastka P, et al. Fecal microbiota transplantation for the improvement of metabolism in obesity: the FMT-TRIM double-blind placebo-controlled pilot trial. PLoS Med 2020;17:e1003051.
127. Leong KSW, Jayasinghe TN, Wilson BC, et al. Effects of fecal microbiome transfer in adolescents with obesity: the gut bugs randomized controlled trial. JAMA Netw Open 2020;3:e2030415.
128. Machado MV. Nonalcoholic fatty liver disease in lean subjects: is it all metabolic-associated fatty liver disease? HR 2020:2020.
129. Lonardo A, Bellentani S, Argo CK, et al. Non-alcoholic Fatty Liver Disease Study Group. Epidemiological modifiers of non-alcoholic fatty liver disease: focus on high-risk groups. Dig Liver Dis 2015;47:997-1006.
130. Le Roy T, Llopis M, Lepage P, et al. Intestinal microbiota determines development of non-alcoholic fatty liver disease in mice. Gut 2013;62:1787-94.
131. Henao-Mejia J, Elinav E, Jin C, et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012;482:179-85.
132. De Minicis S, Rychlicki C, Agostinelli L, et al. Dysbiosis contributes to fibrogenesis in the course of chronic liver injury in mice. Hepatology 2014;59:1738-49.
133. Chiu CC, Ching YH, Li YP, et al. Nonalcoholic fatty liver disease is exacerbated in high-fat diet-fed gnotobiotic mice by colonization with the gut microbiota from patients with nonalcoholic steatohepatitis. Nutrients 2017;9:1220.
134. Hoyles L, Fernández-Real JM, Federici M, et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat Med 2018;24:1070-80.
135. Bergheim I, Weber S, Vos M, et al. Antibiotics protect against fructose-induced hepatic lipid accumulation in mice: role of endotoxin. J Hepatol 2008;48:983-92.
136. Membrez M, Blancher F, Jaquet M, et al. Gut microbiota modulation with norfloxacin and ampicillin enhances glucose tolerance in mice. FASEB J 2008;22:2416-26.
137. Drenick EJ, Fisler J, Johnson D. Hepatic steatosis after intestinal bypass--prevention and reversal by metronidazole, irrespective of protein-calorie malnutrition. Gastroenterology 1982;82:535-48.
138. Wijarnpreecha K, Lou S, Watthanasuntorn K, et al. Small intestinal bacterial overgrowth and nonalcoholic fatty liver disease: a systematic review and meta-analysis. Eur J Gastroenterol Hepatol 2020;32:601-8.
139. Kuang L, Zhou W, Jiang Y. Association of small intestinal bacterial overgrowth with nonalcoholic fatty liver disease in children: a meta-analysis. PLoS One 2021;16:e0260479.
140. Kessoku T, Kobayashi T, Tanaka K, et al. The role of leaky gut in nonalcoholic fatty liver disease: a novel therapeutic target. Int J Mol Sci 2021;22:8161.
141. Luther J, Garber JJ, Khalili H, et al. Hepatic Injury in nonalcoholic steatohepatitis contributes to altered intestinal permeability. Cell Mol Gastroenterol Hepatol 2015;1:222-32.
142. Lang S, Schnabl B. Microbiota and fatty liver disease-the known, the unknown, and the future. Cell Host Microbe 2020;28:233-44.
143. Jiang W, Wu N, Wang X, et al. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci Rep 2015;5:8096.
144. Zhu L, Baker SS, Gill C, et al. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: a connection between endogenous alcohol and NASH. Hepatology 2013;57:601-9.
145. Crovesy L, Masterson D, Rosado EL. Profile of the gut microbiota of adults with obesity: a systematic review. Eur J Clin Nutr 2020;74:1251-62.
146. Del Chierico F, Nobili V, Vernocchi P, et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017;65:451-64.
147. Oh JH, Lee JH, Cho MS, et al. Characterization of gut microbiome in Korean patients with metabolic associated fatty liver disease. Nutrients 2021;13:1013.
148. Mouzaki M, Comelli EM, Arendt BM, et al. Intestinal microbiota in patients with nonalcoholic fatty liver disease. Hepatology 2013;58:120-7.
149. Adams LA, Wang Z, Liddle C, et al. Bile acids associate with specific gut microbiota, low-level alcohol consumption and liver fibrosis in patients with non-alcoholic fatty liver disease. Liver Int 2020;40:1356-65.
150. Da Silva HE, Teterina A, Comelli EM, et al. Nonalcoholic fatty liver disease is associated with dysbiosis independent of body mass index and insulin resistance. Sci Rep 2018;8:1466.
151. Schwimmer JB, Johnson JS, Angeles JE, et al. Microbiome signatures associated with steatohepatitis and moderate to severe fibrosis in children with nonalcoholic fatty liver disease. Gastroenterology 2019;157:1109-22.
152. Wong VW, Tse CH, Lam TT, et al. Molecular characterization of the fecal microbiota in patients with nonalcoholic steatohepatitis--a longitudinal study. PLoS One 2013;8:e62885.
153. Raman M, Ahmed I, Gillevet PM, et al. Fecal microbiome and volatile organic compound metabolome in obese humans with nonalcoholic fatty liver disease. Clin Gastroenterol Hepatol 2013;11:868-75.e1.
154. Li F, Ye J, Shao C, Zhong B. Compositional alterations of gut microbiota in nonalcoholic fatty liver disease patients: a systematic review and Meta-analysis. Lipids Health Dis 2021;20:22.
155. Lee G, You HJ, Bajaj JS, et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat Commun 2020;11:4982.
156. Lay C, Sutren M, Rochet V, Saunier K, Doré J, Rigottier-Gois L. Design and validation of 16S rRNA probes to enumerate members of the Clostridium leptum subgroup in human faecal microbiota. Environ Microbiol 2005;7:933-46.
157. Munukka E, Rintala A, Toivonen R, et al. Faecalibacterium prausnitzii treatment improves hepatic health and reduces adipose tissue inflammation in high-fat fed mice. ISME J 2017;11:1667-79.
158. Miquel S, Leclerc M, Martin R, et al. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. mBio 2015;6:e00300-15.
159. Quévrain E, Maubert MA, Michon C, et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn's disease. Gut 2016;65:415-25.
160. Chen YM, Liu Y, Zhou RF, et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci Rep 2016;6:19076.
161. Koeth RA, Wang Z, Levison BS, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 2013;19:576-85.
162. Zhao G, He F, Wu C, et al. Betaine in inflammation: mechanistic aspects and applications. Front Immunol 2018;9:1070.
163. Cole LK, Vance JE, Vance DE. Phosphatidylcholine biosynthesis and lipoprotein metabolism. Biochim Biophys Acta 2012;1821:754-61.
164. Tan X, Liu Y, Long J, et al. Trimethylamine N-Oxide aggravates liver steatosis through modulation of bile acid metabolism and inhibition of farnesoid X receptor signaling in nonalcoholic fatty liver disease. Mol Nutr Food Res 2019;63:e1900257.
165. Romano KA, Vivas EI, Amador-Noguez D, Rey FE. Intestinal microbiota composition modulates choline bioavailability from diet and accumulation of the proatherogenic metabolite trimethylamine-N-oxide. mBio 2015;6:e02481.
166. Xin FZ, Zhao ZH, Liu XL, et al. Escherichia fergusonii promotes nonobese nonalcoholic fatty liver disease by interfering with host hepatic lipid metabolism through its own msRNA 23487. Cell Mol Gastroenterol Hepatol 2022;13:827-41.
167. Volynets V, Küper MA, Strahl S, et al. Nutrition, intestinal permeability, and blood ethanol levels are altered in patients with nonalcoholic fatty liver disease (NAFLD). Dig Dis Sci 2012;57:1932-41.
168. Dawes E, Foster SM. The formation of ethanol in Escherichia coli. Biochimica et Biophysica Acta 1956;22:253-65.
169. Caussy C, Tripathi A, Humphrey G, et al. A gut microbiome signature for cirrhosis due to nonalcoholic fatty liver disease. Nat Commun 2019;10:1406.
170. Looft T, Johnson TA, Allen HK, et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc Natl Acad Sci U S A 2012;109:1691-6.
171. Yuan J, Chen C, Cui J, et al. Fatty liver disease caused by high-alcohol-producing klebsiella pneumoniae. Cell Metab 2019;30:675-688.e7.
172. Bayoumy AB, Mulder CJJ, Mol JJ, Tushuizen ME. Gut fermentation syndrome: a systematic review of case reports. United European Gastroenterol J 2021;9:332-42.
173. Chen X, Zhang Z, Li H, et al. Endogenous ethanol produced by intestinal bacteria induces mitochondrial dysfunction in non-alcoholic fatty liver disease. J Gastroenterol Hepatol 2020;35:2009-19.
174. McDonnell L, Gilkes A, Ashworth M, et al. Association between antibiotics and gut microbiome dysbiosis in children: systematic review and meta-analysis. Gut Microbes 2021;13:1-18.
175. Lebeer S, Vanderleyden J, De Keersmaecker SC. Genes and molecules of lactobacilli supporting probiotic action. Microbiol Mol Biol Rev 2008;72:728-64, Table of Contents.
176. Elshaghabee FM, Bockelmann W, Meske D, et al. Ethanol production by selected intestinal microorganisms and lactic acid bacteria growing under different nutritional conditions. Front Microbiol 2016;7:47.
177. Boursier J, Mueller O, Barret M, et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016;63:764-75.
178. Dietz FK, Traiger GJ. Potentiation of CCl4 of hepatotoxicity in rats by a metabolite of 2-butanone: 2,3-butanediol. Toxicology 1979;14:209-15.
179. Hewitt L. Modifications in rat hepatobiliary function following treatment with acetone, 2-butanone, 2-hexanone, mirex, or chlordecone and subsequently exposed to chloroform*1, *2. Toxicology and Applied Pharmacology 1986;83:465-73.
180. Urdaneta V, Casadesús J. Interactions between bacteria and bile salts in the gastrointestinal and hepatobiliary tracts. Front Med (Lausanne) 2017;4:163.
181. Machado MV, Cortez-Pinto H. Nuclear receptors: how do they position in non-alcoholic fatty liver disease treatment? Liver Int 2014;34:1291-4.
182. Wang B, Jiang X, Cao M, et al. Altered fecal microbiota correlates with liver biochemistry in nonobese patients with non-alcoholic fatty liver disease. Sci Rep 2016;6:32002.
183. Duarte SMB, Stefano JT, Miele L, et al. Gut microbiome composition in lean patients with NASH is associated with liver damage independent of caloric intake: a prospective pilot study. Nutr Metab Cardiovasc Dis 2018;28:369-84.
184. Demir M, Lang S, Hartmann P, et al. The fecal mycobiome in non-alcoholic fatty liver disease. J Hepatol 2022;76:788-99.
185. Fotis D, Liu J, Dalamaga M. Could gut mycobiome play a role in NAFLD pathogenesis? Metabol Open 2022;14:100178.
186. Taylor RS, Taylor RJ, Bayliss S, et al. Association between fibrosis stage and outcomes of patients with nonalcoholic fatty liver disease: a systematic review and meta-analysis. Gastroenterology 2020;158:1611-1625.e12.
187. Loomba R, Seguritan V, Li W, et al. Gut microbiome-based metagenomic signature for non-invasive detection of advanced fibrosis in human nonalcoholic fatty liver disease. Cell Metab 2017;25:1054-1062.e5.
188. Oh TG, Kim SM, Caussy C, et al. A universal gut-microbiome-derived signature predicts cirrhosis. Cell Metab 2020;32:878-888.e6.
189. Lanthier N, Rodriguez J, Nachit M, et al. Microbiota analysis and transient elastography reveal new extra-hepatic components of liver steatosis and fibrosis in obese patients. Sci Rep 2021;11:659.
190. Kwan SY, Jiao J, Joon A, et al. Gut microbiome features associated with liver fibrosis in Hispanics, a population at high risk for fatty liver disease. Hepatology 2022;75:955-67.
191. Dong TS, Katzka W, Lagishetty V, et al. A microbial signature identifies advanced fibrosis in patients with chronic liver disease mainly due to NAFLD. Sci Rep 2020;10:2771.
193. Kaakoush NO. Insights into the role of erysipelotrichaceae in the human host. Front Cell Infect Microbiol 2015;5:84.
194. Aron-Wisnewsky J, Prifti E, Belda E, et al. Major microbiota dysbiosis in severe obesity: fate after bariatric surgery. Gut 2019;68:70-82.
195. Kummen M, Thingholm LB, Rühlemann MC, et al. Altered gut microbial metabolism of essential nutrients in primary sclerosing cholangitis. Gastroenterology 2021;160:1784-1798.e0.
196. Visekruna A, Luu M. The role of short-chain fatty acids and bile acids in intestinal and liver function, inflammation, and carcinogenesis. Front Cell Dev Biol 2021;9:703218.
197. Canivet CM, David N, Pailhoriès H, et al. Cross-linkage between bacterial taxonomy and gene functions: a study of metagenome-assembled genomes of gut microbiota in adult non-alcoholic fatty liver disease. Aliment Pharmacol Ther 2021;53:722-32.
198. Lang S, Demir M, Martin A, et al. Intestinal virome signature associated with severity of nonalcoholic fatty liver disease. Gastroenterology 2020;159:1839-52.
199. Qin N, Yang F, Li A, et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014;513:59-64.
200. Ponziani FR, Bhoori S, Castelli C, et al. Hepatocellular carcinoma is associated with gut microbiota profile and inflammation in nonalcoholic fatty liver disease. Hepatology 2019;69:107-20.
201. Bajaj JS, Betrapally NS, Hylemon PB, et al. Gut microbiota alterations can predict hospitalizations in cirrhosis independent of diabetes mellitus. Sci Rep 2015;5:18559.
202. Chen Y, Guo J, Qian G, et al. Gut dysbiosis in acute-on-chronic liver failure and its predictive value for mortality. J Gastroenterol Hepatol 2015;30:1429-37.
203. Bajaj JS, Heuman DM, Hylemon PB, et al. Altered profile of human gut microbiome is associated with cirrhosis and its complications. J Hepatol 2014;60:940-7.
204. Behary J, Amorim N, Jiang XT, et al. Gut microbiota impact on the peripheral immune response in non-alcoholic fatty liver disease related hepatocellular carcinoma. Nat Commun 2021;12:187.
205. Sydor S, Best J, Messerschmidt I, et al. Altered microbiota diversity and bile acid signaling in cirrhotic and noncirrhotic NASH-HCC. Clin Transl Gastroenterol 2020;11:e00131.
206. Yoshimoto S, Loo TM, Atarashi K, et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature 2013;499:97-101.
207. Ma C, Han M, Heinrich B, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science 2018;360:eaan5931.
208. Wu H, Esteve E, Tremaroli V, et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat Med 2017;23:850-8.
209. Moreira GV, Azevedo FF, Ribeiro LM, et al. Liraglutide modulates gut microbiota and reduces NAFLD in obese mice. J Nutr Biochem 2018;62:143-54.
210. Maier L, Pruteanu M, Kuhn M, et al. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018;555:623-8.
211. Sun L, Xie C, Wang G, et al. Gut microbiota and intestinal FXR mediate the clinical benefits of metformin. Nat Med 2018;24:1919-29.
212. Lee H, Ko G. Effect of metformin on metabolic improvement and gut microbiota. Appl Environ Microbiol 2014;80:5935-43.
213. Zhang S, Wong YT, Tang KY, Kwan HY, Su T. Chinese medicinal herbs targeting the gut-liver axis and adipose tissue-liver axis for non-alcoholic fatty liver disease treatments: the ancient wisdom and modern science. Front Endocrinol (Lausanne) 2020;11:572729.
214. Wang RR, Zhang LF, Chen LP, et al. Structural and functional modulation of gut microbiota by jiangzhi granules during the amelioration of nonalcoholic fatty liver disease. Oxid Med Cell Longev 2021;2021:2234695.
215. Jin W, Cho S, Laxi N, et al. Hepatoprotective Effects of ixeris chinensis on nonalcoholic fatty liver disease induced by high-fat diet in mice: an integrated gut microbiota and metabolomic analysis. Molecules 2022;27:3148.
216. Lang S, Martin A, Farowski F, et al. High protein intake is associated with histological disease activity in patients with NAFLD. Hepatol Commun 2020;4:681-95.
217. Martinez JE, Kahana DD, Ghuman S, et al. Unhealthy lifestyle and gut dysbiosis: a better understanding of the effects of poor diet and nicotine on the intestinal microbiome. Front Endocrinol (Lausanne) 2021;12:667066.
218. Khan MY, Mihali AB, Rawala MS, Aslam A, Siddiqui WJ. The promising role of probiotic and synbiotic therapy in aminotransferase levels and inflammatory markers in patients with nonalcoholic fatty liver disease - a systematic review and meta-analysis. Eur J Gastroenterol Hepatol 2019;31:703-15.
219. Sharpton SR, Maraj B, Harding-Theobald E, Vittinghoff E, Terrault NA. Gut microbiome-targeted therapies in nonalcoholic fatty liver disease: a systematic review, meta-analysis, and meta-regression. Am J Clin Nutr 2019;110:139-49.
220. Yang R, Shang J, Zhou Y, Liu W, Tian Y, Shang H. Effects of probiotics on nonalcoholic fatty liver disease: a systematic review and meta-analysis. Expert Rev Gastroenterol Hepatol 2021;15:1401-9.
221. Kristensen NB, Bryrup T, Allin KH, Nielsen T, Hansen TH, Pedersen O. Alterations in fecal microbiota composition by probiotic supplementation in healthy adults: a systematic review of randomized controlled trials. Genome Med 2016:8.
222. Maldonado-Gómez MX, Martínez I, Bottacini F, et al. Stable engraftment of bifidobacterium longum AH1206 in the human gut depends on individualized features of the resident microbiome. Cell Host Microbe 2016;20:515-26.
223. Loman BR, Hernández-Saavedra D, An R, Rector RS. Prebiotic and probiotic treatment of nonalcoholic fatty liver disease: a systematic review and meta-analysis. Nutr Rev 2018;76:822-39.
224. Gangarapu V, Ince AT, Baysal B, et al. Efficacy of rifaximin on circulating endotoxins and cytokines in patients with nonalcoholic fatty liver disease. Eur J Gastroenterol Hepatol 2015;27:840-5.
225. Abdel-Razik A, Mousa N, Shabana W, et al. Rifaximin in nonalcoholic fatty liver disease: hit multiple targets with a single shot. Eur J Gastroenterol Hepatol 2018;30:1237-46.
226. Cobbold JFL, Atkinson S, Marchesi JR, et al. Rifaximin in non-alcoholic steatohepatitis: an open-label pilot study. Hepatol Res 2018;48:69-77.
227. Proença IM, Allegretti JR, Bernardo WM, et al. Fecal microbiota transplantation improves metabolic syndrome parameters: systematic review with meta-analysis based on randomized clinical trials. Nutr Res 2020;83:1-14.
228. Ng SC, Xu Z, Mak JWY, et al. Microbiota engraftment after faecal microbiota transplantation in obese subjects with type 2 diabetes: a 24-week, double-blind, randomised controlled trial. Gut 2022;71:716-23.
229. Craven L, Rahman A, Nair Parvathy S, et al. Allogenic fecal microbiota transplantation in patients with nonalcoholic fatty liver disease improves abnormal small intestinal permeability: a randomized control trial. Am J Gastroenterol 2020;115:1055-65.
230. Sharpton SR, Schnabl B, Knight R, Loomba R. Current concepts, opportunities, and challenges of gut microbiome-based personalized medicine in nonalcoholic fatty liver disease. Cell Metab 2021;33:21-32.
Cite This Article
Export citation file: BibTeX | RIS
OAE Style
Sousa P, Machado MV. MAFLD under the lens: the role of gut microbiota. Metab Target Organ Damage 2022;2:14. http://dx.doi.org/10.20517/mtod.2022.15
AMA Style
Sousa P, Machado MV. MAFLD under the lens: the role of gut microbiota. Metabolism and Target Organ Damage. 2022; 2(4): 14. http://dx.doi.org/10.20517/mtod.2022.15
Chicago/Turabian Style
Sousa, Patrícia, Mariana Verdelho Machado. 2022. "MAFLD under the lens: the role of gut microbiota" Metabolism and Target Organ Damage. 2, no.4: 14. http://dx.doi.org/10.20517/mtod.2022.15
ACS Style
Sousa, P.; Machado MV. MAFLD under the lens: the role of gut microbiota. Metab Target Organ Damage. 2022, 2, 14. http://dx.doi.org/10.20517/mtod.2022.15
About This Article
Copyright

Data & Comments
Data


 Cite This Article 16 clicks
Cite This Article 16 clicks

 Like This Article 3
likes
Like This Article 3
likes







Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.